 東京大学 研究
東京大学 研究世界初!葉緑体ゲノム編集でRubiscoを強化し光合成と植物生産性の向上に成功
――光合成を設計する次世代植物育種技術への期待――
【注目の成果:共同研究・産学連携のためのチェックポイント】
 | 外来遺伝子をゲノムに残さない、精密な葉緑体ゲノム編集によって光合成能力と作物生産性を同時に向上させ得ることを示した点で画期的 |
【産学連携対象 全学共通分野 Discovery Saga】
【Sagaキーワード】
先端技術/DNA結合/産学連携/光エネルギー/人口増加/気候変動/二量体/触媒反応/ロイシン/クロロフィル/コドン/光合成/葉緑体/カーボンニュートラル/持続可能/反応速度/カーボン/生産性/電子顕微鏡/二酸化炭素/有機物/CO2固定/遺伝子改変/T-DNA/トウモロコシ/高CO2/変異体/シロイヌナズナ/トマト/イネ/ゲノム編集技術/CO2濃度/バイオマス/水利用/クライオ電子顕微鏡/将来予測/分子機構/アミノ酸置換/ゲノム編集/点変異/RNA/アミノ酸/ラット/遺伝子導入/核酸塩基/細胞核/創薬/ゲノム/遺伝子
発表のポイント
◆Rubiscoは光合成における最も重要な酵素ですが、触媒効率が悪いため、Rubiscoの機能を上げることが作物の生産性向上につながります。しかし、Rubiscoの触媒に関わる部分(Rubisco大サブユニット)の遺伝子は、細胞核のゲノムとは別の葉緑体ゲノムにコードされているため、その精密な改変はこれまで技術的に困難でした。◆本研究では、葉緑体ゲノム編集によりRubisco大サブユニットに単一アミノ酸置換を導入し、触媒速度(kcat)を上げることで、現在および将来予測される高CO₂条件下での光合成能力と作物生産性の向上に成功しました。
◆本研究は、外来遺伝子をゲノムに残さない精密な葉緑体ゲノム編集により、光合成能力と作物生産性を向上させ、大気CO₂削減への貢献も期待される新たな分子育種戦略の可能性を示すものです。
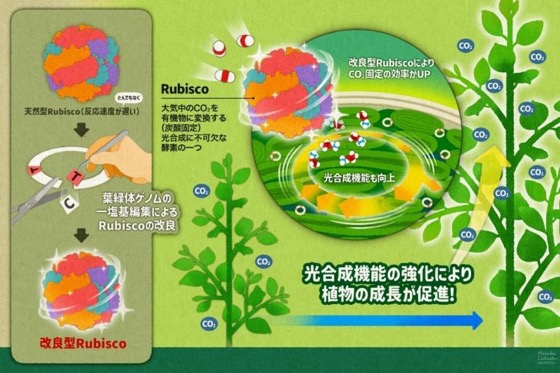 本研究のイメージ図
本研究のイメージ図葉緑体ゲノム編集による高機能Rubiscoの創出で光合成と植物生産性の向上に成功© ウチダヒロコ
発表概要
東京大学大学院農学生命科学研究科の矢守航准教授、中里一星特任助教、有村慎一教授、立命館大学の松村浩由教授、神戸大学の深山浩教授、大阪大学の難波啓一特任教授らの研究グループは、世界的な食料需要の増加と気候変動への対応に向けて、植物の光合成能力を分子レベルで強化することに成功しました。光合成において炭素固定を担う中心酵素Rubiscoは、その低い触媒効率が作物生産性を制限する要因として長年知られてきましたが、触媒部位を構成するRubisco大サブユニットは細胞核のゲノムとは異なる葉緑体ゲノムにコードされているため、その機能を精密に改変することは極めて困難とされてきました。本研究では、葉緑体ゲノム編集技術を用いてRubisco大サブユニット遺伝子(rbcL)に単一アミノ酸置換を導入し、酵素量を維持したまま触媒速度(kcat)を向上させることに世界に先駆けて成功しました。その結果、現在の大気CO₂濃度条件に加え、将来予測される高CO₂環境においてもCO₂同化速度の向上と植物バイオマスの増加が確認され、光合成能力の分子改良が実際の植物成長や生産性向上につながることが明確に示されました。さらにクライオ電子顕微鏡解析により、Rubiscoのkcatが上がった原因を調べたところ、活性中心から離れた位置の単一アミノ酸置換が触媒部位の柔軟性を変化させ、酵素機能を高める新たな分子機構の可能性が示されました。
本成果は、外来遺伝子をゲノムに残さない、精密な葉緑体ゲノム編集によって光合成能力と作物生産性を同時に向上させ得ることを示した点で画期的であり、作物の進化や育種を加速する新たな分子育種戦略の確立につながるものです。今後、主要作物や樹木への展開により、食料生産性の向上に加えて光合成による炭素固定能力の強化を通じた大気CO₂削減やカーボンニュートラル社会の実現への貢献が期待されます。
背景
気候変動の進行と人口増加により、世界の食料生産は2050年までに30%以上の増加が必要とされています。作物生産性を根本的に規定する要因の一つが光合成能力であり、その効率向上は持続可能な農業の実現に向けた最重要課題とされています。光合成においてCO₂固定を担う中心酵素Rubiscoは、触媒効率が低く、さらに酸素化反応を併発するという特性から光合成の律速要因となっています。シロイヌナズナやイネ、コムギなどの主要な農作物はC3植物(注1)に属しますが、これらC3植物のRubiscoは触媒速度が比較的低いことが知られています。一方で、トウモロコシやソルガムなどのC4植物(注1)は、触媒速度(kcat)が高いRubiscoを有しており、高い光合成能力を示します。モデル解析の結果からも、C4植物型のRubisco特性をC3植物に導入できれば、現在の大気CO₂条件でも光合成速度が20~50%向上する可能性があると報告されています。
将来的に大気中のCO₂濃度の上昇が予測される中では、このように反応速度(kcat)が高いRubiscoがより効率的に機能すると考えられています。そこで本研究では、C3植物のRubiscoの触媒特性を、光合成能力の高いC4植物のRubiscoに近づけること、すなわち「RubiscoのC4化」を目指しました。
しかし、これまでにも「高機能Rubisco」の導入が試みられてきたものの、異種酵素の導入に伴う酵素組立の不全や成長低下などの問題があり、実際に生産性の向上につながる例は限られていました。加えて、触媒部位を担うRubisco大サブユニットは葉緑体ゲノムにコードされているため、その機能を精密に改変すること自体が技術的に大きな課題とされてきました。
そこで本研究では、C3植物のモデル植物であるシロイヌナズナを用い、他種の遺伝子導入に依存せず既存のRubisco配列を最小限に改変する葉緑体ゲノム編集という新たな光合成改良戦略に着目しました。
発表内容
本研究では、標的一塩基置換酵素(ptpTALECD)(注2)を用いて、C3植物のモデル植物であるシロイヌナズナの葉緑体ゲノムにコードされる光合成酵素Rubiscoの大サブユニット遺伝子(rbcL)に精密な点変異を導入しました。具体的には、309番目のアミノ酸メチオニンをイソロイシンに置換した変異体(M309I)や、397番目のアミノ酸アスパラギン酸をアスパラギンに置換した変異体(D397N)を含む、合計7種類の単一アミノ酸改変系統を作製しました(図1)。標的としたアミノ酸の位置はRubiscoの三次元構造上に示しており、触媒反応が行われる活性中心との位置関係が分かるようにしています(図1a)。また、導入を試みた置換変異、対応するコドン(注3)の変化、および実際に得られた変異体の遺伝子型を整理して示しました(図1b)。
次に、各改変系統におけるRubiscoの酵素特性を解析しました(図2)。その結果、多くの改変系統では触媒性能の向上は見られず、光合成能力や生育が低下する場合も確認され、Rubisco機能改良の難しさが改めて示されました。一方で、M309IおよびD397N変異体では触媒速度(kcat)が有意に向上していました(図2a)。CO₂親和性の指標であるKc(値が低いほどCO₂に結合しやすい)は、M309Iでは上昇し、D397Nでは野生型(Col-0)と同程度でした(図2b)。その結果、CO₂固定効率の指標であるkcat/KcはM309Iでは低下し、D397Nでは維持されました(図2c)。また、CO₂固定と酸素反応の比を示すSc/oは両変異体で低下しました(図2d)。これらの特徴は、一般に高CO₂環境で効率よく機能するC4植物型Rubiscoの特性と共通しており、本研究によりC3植物Rubiscoの触媒特性がC4化されたことを示します。これらの変化はRubisco量(図2e)やクロロフィル(注4)の量(図2f)の変化を伴わずに生じていました。
この酵素性能の変化は植物個体の光合成機能にも直接反映されました。現在の大気CO₂濃度条件下において、改変系統ではCO₂同化速度が増加しました(図3a)。一方で、気孔コンダクタンス(図3b、注5)や葉内CO₂濃度(図3c)には差が見られず、その結果として水利用効率が向上しました(図3d)。これはRubisco機能そのものの強化によって光合成効率が高まったことを示しています。
さらに生育解析の結果、播種48日後には現在の大気CO₂条件下において植物の外観からも生育促進が認められ(図4a)、総葉面積(図4b)および地上部乾物重(図4c)が増加していました。将来の気候環境を想定した高CO₂条件下ではこの効果はさらに顕著となり、植物の外観(図4d)、総葉面積(図4e)、地上部乾物重(図4f)がより大きく増加しました。
最終的な生産性評価として、播種90日後に収穫した植物では、生育外観の改善(図5a)、地上部乾物重(図5b)および地下部乾物重(図5c)の増加に加え、種子収量も増加していました(図5d)。これは光合成能力の向上が最終的な収量増加につながることを示しています。
さらにクライオ電子顕微鏡によって、Col-0、M309I、D397Nの構造解析を行いました。活性部位の構造に着目すると、Col-0では基質アナログ(注6)である硫酸イオンが2か所に結合しているのに対し(図6b)、M309IおよびD397Nでは硫酸イオンが1か所のみに結合していました(図6c–f)。さらに、RubiscoがCO₂を固定する際に開閉を伴う60sループの構造にも変化が確認されました(図6d, f)。アミノ酸変異を導入したM309IやD397Nの位置は活性中心から離れたサブユニット界面に位置しています(図6a)。つまり、活性部位から離れた位置の単一アミノ酸置換によって、ドミノ倒しのように酵素の構造が変化し、それによって触媒部位の柔軟性が高まったことでRubiscoの触媒速度が向上した可能性が分かりました。
以上の結果は、ゲノム編集技術を用いて、C3植物におけるRubiscoの触媒特性をC4型に近づけることで、光合成能力と植物生産性を同時に向上できることを示した重要な成果です。
今後の展開と意義
本研究は、葉緑体ゲノム編集技術を用いてRubiscoの触媒特性を単一アミノ酸レベルという最小限の改変によって向上させ、光合成能力と植物生産性を同時に強化できることを世界に先駆けて示した重要な成果です。葉緑体ゲノム編集は、外来遺伝子をゲノムに残すことなく精密な遺伝子改変が可能な技術であり、従来の遺伝子組換えとは異なる形で植物の進化や育種のスピードを飛躍的に高め得る新しい分子育種手法としての可能性が本研究で示されました。今後、本技術をイネやトマトなどの主要作物やポプラなどの木本植物へ展開することで、作物生産性の大幅な向上に加え、光合成による炭素固定能力の強化を通じた大気CO₂削減やカーボンニュートラル社会の実現に貢献する次世代の光合成改良技術としての発展が強く期待されます。
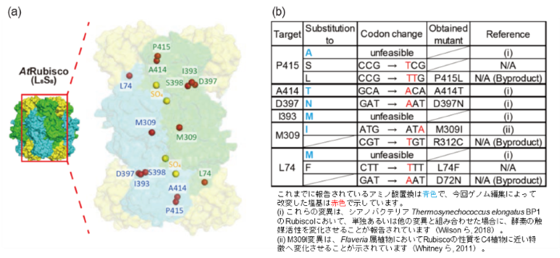 図1. RbcLに導入したアミノ酸置換
図1. RbcLに導入したアミノ酸置換
a:標的としたアミノ酸の位置を、シロイヌナズナRubisco(AtRubisco)の三次元構造上に示した。RbcL二量体および4つのRbcSサブユニットは半透明の表面モデルで表し、それぞれ緑色、青色、黄色で示している。触媒部位に結合した硫酸分子は、触媒中心を示すため黄色の球で示した。標的アミノ酸のCα原子の位置は赤色の球で示した。
b:標的アミノ酸、導入を目指した置換変異、それに対応するコドン変化、さらに得られた変異体およびその遺伝子型を示した。「unfeasible」は、C:GからT:Aへの塩基編集では導入できない置換であることを示す。ホモプラスミーはT2世代個体およびそのT3後代の遺伝子型解析によって確認した。NullはT-DNAが存在しないことを示す。
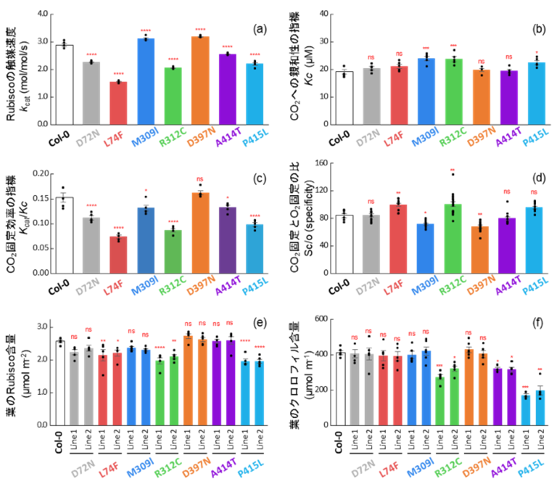 図2. 各変異系統におけるRubiscoの酵素特性、Rubisco量およびクロロフィル量
図2. 各変異系統におけるRubiscoの酵素特性、Rubisco量およびクロロフィル量
各種変異を導入したRubiscoの酵素特性として、触媒速度(kcat)(a)、CO₂に対する親和性の指標(Kc)(b)、CO₂固定効率(kcat/Kc)(c)、CO₂固定と酸素反応の比(Sc/o)(d)、Rubisco量(e)、およびクロロフィル量(f)を示す。
データは平均値±標準誤差(SE)で示し、黒点は各測定値を示す。ns は有意差なし(p > 0.05)、* p< 0.05、** p< 0.01、*** p< 0.001、**** p< 0.0001 を示す。独立した酵素調製6回の結果に基づき、Dunnettの両側検定による多重比較を行った。
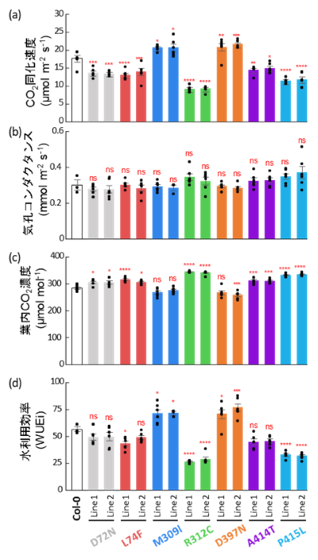 図3. 各系統における光合成関連パラメータ
図3. 各系統における光合成関連パラメータ
大気CO₂濃度400 ppm条件下における各系統のCO₂同化速度(a)、気孔コンダクタンス(b)、葉内CO₂濃度(c)、および固有水利用効率(WUEi)(d)を示す。
データは平均値±標準誤差(SE)で示し、黒点は個々の測定値を示す。ns は有意差なし(p > 0.05)、* p< 0.05、** p< 0.01、*** p< 0.001、**** p< 0.0001 を示す。各系統6個体の測定結果に基づき、Dunnettの両側検定による多重比較を行った。
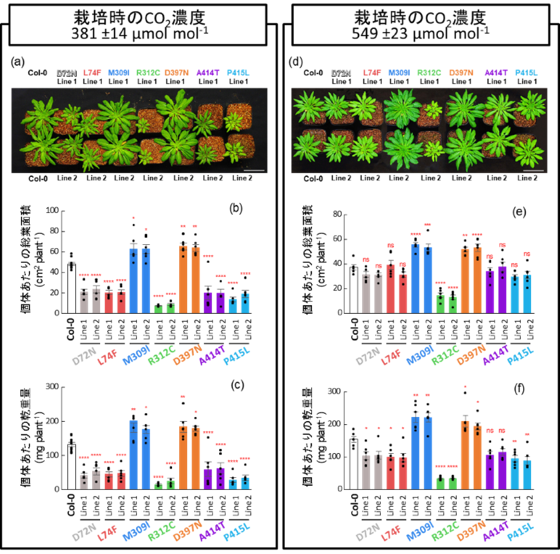 図4. 播種後48日における植物の生育
図4. 播種後48日における植物の生育
現在の大気CO₂濃度条件(381 ±14 µmol mol⁻¹)で栽培した植物の外観(a)、播種48日後における総葉面積(b)および地上部乾物重(c)を示す。高CO₂条件(549 ±23 µmol mol⁻¹)で栽培した植物の外観(d)、総葉面積(e)および地上部乾物重(f)を示す。
データは平均値±標準誤差(SE)で示し、黒点は個々の測定値を示す。ns は有意差なし(p > 0.05)、* p< 0.05、** p< 0.01、*** p< 0.001、**** p< 0.0001 を示す。各系統6個体の測定結果に基づき、Dunnettの両側検定による多重比較を行った。
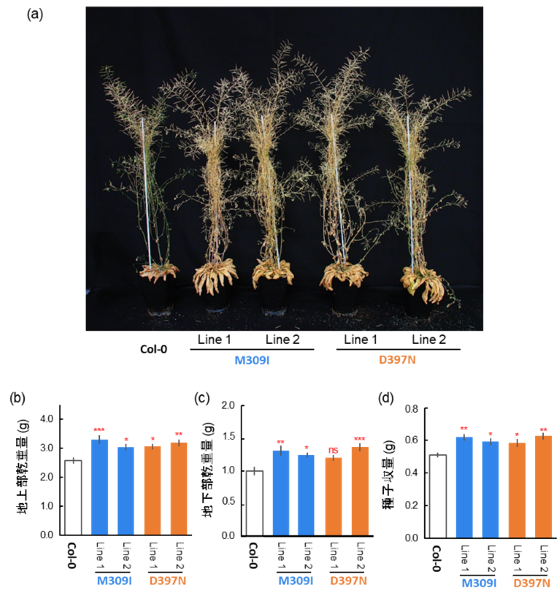 図5. 現在の大気CO₂濃度条件(381±14 μmol mol⁻¹)における植物の生育
図5. 現在の大気CO₂濃度条件(381±14 μmol mol⁻¹)における植物の生育
播種90日後に収穫した植物の外観(a)、地上部乾物重(b)、地下部乾物重(c)および種子収量(d)を示す。
植物は明期/暗期10/14時間の条件で栽培し、明期の光合成光量子束密度(PPFD)は150 μmol photons m⁻² s⁻¹に設定した。気温および相対湿度はそれぞれ22℃および60%に維持した。
データは平均値±標準誤差(SE)(n=5)で示す。ns は有意差なし(p > 0.05)、* p< 0.05、** p< 0.01、*** p< 0.001 を示す。各列はCol-0と比較した有意差を示し、多重比較にはDunnettの両側検定を用いた。
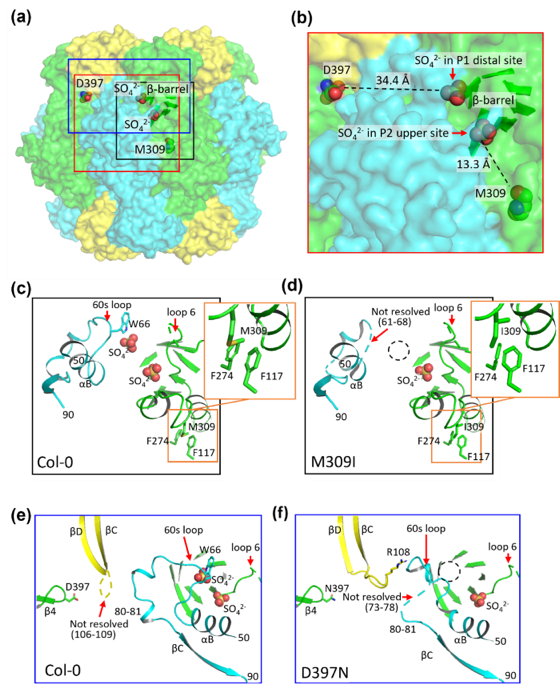 図6. クライオ電子顕微鏡による各系統Rubiscoの構造解析
図6. クライオ電子顕微鏡による各系統Rubiscoの構造解析
a:野生型(Col-0)RubiscoにおけるM309およびD397の位置を示す。8つのRbcLサブユニットは緑色およびシアン色、8つのRbcSサブユニットは黄色で示している。M309およびD397、ならびに2つの硫酸イオン(SO42-)は空間充填モデルで表示した。活性部位を形成するC末端領域を含むβバレル構造も示している。赤、黒、青の四角は、それぞれb、c・d、e・fで拡大して示した領域を示す。
b:活性部位周辺の拡大構造。P2上位部位に結合した硫酸イオンとM309との距離、およびP1遠位部位に結合した硫酸イオンとD397との距離を示す。
c、d:Col-0とM309I変異体における活性部位構造の比較。主要な相互作用残基を示している。破線の円はP1遠位部位を示し、この部位では硫酸イオンが観察されなかった。
e、f:Col-0とD397N変異体における活性部位構造の比較。主要な相互作用残基を示している。破線の円はP1遠位部位を示し、この部位では硫酸イオンが観察されなかった。
発表者・研究者等情報
東京大学 大学院農学生命科学研究科矢守 航 准教授
中里 一星 特任助教
曲 玉辰 特任助教
三箇 雪花 修士課程大学院生 (研究当時)
有村 慎一 教授
立命館大学 生命科学部
上原 了 助教
能登 悠真 修士課程大学院生
松村 浩由 教授
神戸大学 大学院農学研究科
深山 浩 教授
大阪大学大学院生命機能研究科 日本電子YOKOGUSHI協働研究所
宮田 知子 特任准教授(常勤)
難波 啓一 特任教授(常勤)
論文情報
雑誌名:Nature Communications題名:Chloroplast genome editing of Rubisco boosts photosynthesis and plant growth
著者名:Wataru Yamori, Issei Nakazato, Yuchen Qu, Yukina Sanga, Tomoko Miyata, Ryo Uehara, Yuma Noto, Keiichi Namba, Hiroshi Fukayama, Hiroyoshi Matsumura, and Shin-ichi Arimura
DOI:10.1038/s41467-026-73783-w
URL:https://www.nature.com/articles/s41467-026-73783-w
研究助成
本研究は、以下の支援を受けて実施されました。日本学術振興会 科学研究費助成事業(課題番号:21H02171、24H02271、24H02277、25H02292、24K01994、23K18033、JPJSCCA20230008、22J20237)、科学技術振興機構(JST)ALCA-Next(課題番号 JPMJAN25D2)、科学技術振興機構(JST)OPERA(産学共創基盤研究プログラム、課題番号 JPMJOP1861)、立命館大学 立命館グローバル・イノベーション研究機構(R-GIRO)研究プログラム、白石科学振興財団、日本医療研究開発機構(AMED)BINDS(創薬等先端技術支援基盤プラットフォーム事業、課題番号 JP21am0101117、JP22ama121003、JP23ama121001)、日本医療研究開発機構(AMED)CiCLE(産学連携医療イノベーション創出プログラム、課題番号 JP17pc0101020);日本電子(JEOL)YOKOGUSHI協働研究所、大阪大学 蛋白質研究所 共同研究プログラム(CR-22-02、CR-23-02)。用語解説
注1. C3植物、C4植物CO2が植物体内に取り込まれた後、最初の反応で炭素を3つ含む有機物に変換される植物がC3植物、炭素を4つ含む有機物に変換される植物がC4植物と呼ばれる。C3植物では植物体内に取り込まれたCO2がRubiscoと最初に反応するのに対し、C4植物ではCO2が別の酵素と反応して有機物に変換された後、別の組織に移動してからCO2へと再変換され、そこでRubiscoと反応する。このように、C3植物とC4植物ではCO2がRubiscoと反応するまでの道のりが異なり、それゆえにRubiscoの性質も異なると考えられている。
注2. 標的一塩基置換酵素ptpTALECD
ptpTALECDはplastid-targeted platinum TALE cytidine deaminaseの略で、DNA結合ドメイン(TALE)と塩基置換を担うドメイン(cytidine deaminase)等から成る、二分子型の塩基置換酵素である。葉緑体ゲノム上の任意のDNA配列に両分子のTALEが結合すると、その間の配列に位置する特定のC:G対がT:A対に置換される。
注3. コドン
タンパク質の設計図となるDNAやRNAにおいて、次にどのアミノ酸をタンパク質に繋げるかを指定する、3つの核酸塩基の並び。
注4. クロロフィル
光合成において光エネルギーを吸収する色素。クロロフィル量は、光合成能力の指標の一つである。
注5. 気孔コンダクタンス
葉の気孔がどれだけ開いているかを表す指標。この値が大きいほど気孔の開き具合が大きく、CO2等の気体が通りやすいことを表す。
注6. 基質アナログ
酵素が本来反応する分子(基質)と似た構造を持ち、酵素の反応部位に結合するが、通常の反応は起こらない化合物。酵素の働きを調べる研究などに用いられる。
問い合わせ先
<研究内容については発表者にお問合せください>東京大学 大学院農学生命科学研究科 附属生態調和農学機構
准教授 矢守 航(やもり わたる)
TEL:070-6442-9511
E-mail:yamori@g.ecc.u-tokyo.ac.jp
東京大学 大学院農学生命科学研究科 生産・環境生物学専攻
教授 有村 慎一(ありむら しんいち)
E-mail:arimura@g.ecc.u-tokyo.ac.jp
東京大学 大学院農学生命科学研究科・農学部
事務部 総務課総務チーム広報情報担当
TEL:03-5841-5484 FAX:03-5841-5028
E-mail:koho.a@gs.mail.u-tokyo.ac.jp
関連教員
矢守 航有村 慎一