 東京大学 研究
東京大学 研究未来の高CO₂環境で「光のゆらぎ」に負けないイネへ
――Rubisco activaseによる光合成と成長の強化戦略――
【注目の成果:共同研究・産学連携のためのチェックポイント】
 | 自然環境を模擬した変動光条件下で、Rubisco activaseの改変によってイネの成長が促進されることを示し、将来の気候変動環境に適応した高生産性作物開発への応用が期待 |
【産学連携対象 全学共通分野 Discovery Saga】
【Sagaキーワード】
プロファイル/人口増加/気候変動/太陽/光化学/クロロフィル/光化学系I/光化学系II/光合成/光阻害/電子伝達/変動光/光環境/太陽光/高温環境/生産性/二酸化炭素/CO2固定/SEM/クロロフィル蛍光/高CO2/水田/イネ/CO2濃度/水利用/ゆらぎ/将来予測/ATP/ゲノム/ストレス
発表のポイント
◆ 光合成酵素Rubiscoを活性化する「Rubisco activase」を増やしたイネでは、変動光環境下で光合成の立ち上がり(光合成誘導)が高速化されることを明らかにしました。◆ 将来予測される高CO₂環境(800 ppm)においても、Rubisco activaseを増やしたイネでは、高い光合成能力と水利用効率が維持されました。
◆ 自然環境を模擬した変動光条件下で、Rubisco activaseの改変によってイネの成長が促進されることを示し、将来の気候変動環境に適応した高生産性作物開発への応用が期待されます。
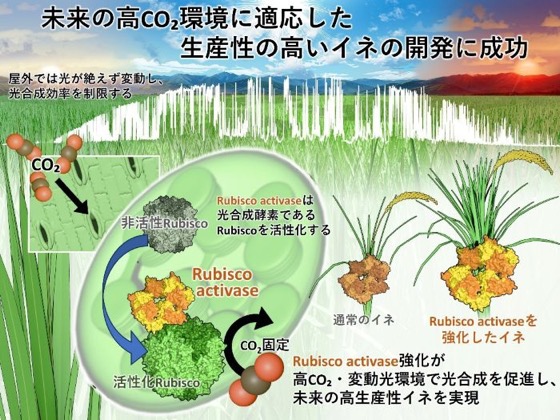 Rubisco activaseによる光合成と成長の強化戦略
Rubisco activaseによる光合成と成長の強化戦略
発表概要
東京大学大学院農学生命科学研究科の矢守 航 准教授らの研究グループは、神戸大学大学院農学研究科の深山 浩 教授との共同研究により、光合成酵素Rubisco(注1)を活性化するタンパク質「Rubisco activase」(注2)を増加させたイネを用いて、将来の高CO₂環境下における変動光環境(注3)への光合成応答を解析しました。自然環境では、植物は雲の移動や葉の揺れ、周囲の葉による遮蔽などによって、秒〜分単位で絶えず変化する光環境にさらされています。このような「変動光環境」では、光が急に強くなっても、最大速度で光合成を行えるようになるまでの「光合成誘導」(注4)に時間がかかることによって、炭素固定量が大きく低下することが知られています。そのため、変動光環境に迅速に応答できる光合成制御機構の解明は、将来の作物生産性向上に向けた重要課題となっています。
本研究では、Rubisco activaseを増やしたイネにおいて、光が弱い状態から強くなった際のRubisco活性化速度が向上し、その結果、光合成誘導が高速化されることを明らかにしました。さらに、将来予測される高CO₂条件(800 ppm)においても、変動光環境下で高い光合成能力を維持し、水利用効率も向上することが分かりました。また、実際の自然環境を模擬した長期的な変動光条件下では、Rubisco activaseを増やしたイネで成長促進も確認されました。
本成果は、変動する光環境への適応能力を高めることで、将来の高CO₂・気候変動環境下でも高い生産性を維持できる作物開発につながる重要な知見です。特に、Rubisco activaseを標的とした光合成改良が、今後の食料生産や気候変動適応型農業に向けた新たな戦略となることが期待されます。
発表内容
植物の光合成は、作物の成長や収量を決定する最も重要な生命活動の一つです。近年、世界的な人口増加や気候変動の進行により、限られた資源でより多くの食料を安定的に生産する技術の開発が求められています。そのため、光合成能力を向上させることは、次世代農業における重要課題として注目されています。一方、植物が実際に生育する自然環境では、葉に当たる光は一定ではありません。雲の移動によって地上に届く太陽光の量が変化したり、風で葉が揺れて光を受ける向きが変わったり、周囲の揺れ動く葉などによって光が遮られたりして、葉が受ける光強度は秒〜分単位が絶えず変動する「変動光環境」です。植物は、光が一旦弱くなって光合成速度が落ちてしまうと、その後急激に光が強くなっても、すぐには最大速度で光合成を行うことができません。この立ち上がりの遅れは「光合成誘導」と呼ばれ、日積算の炭素固定量や作物生産性を大きく制限していると考えられています。
さらに、現在の大気中CO₂濃度は上昇を続けており、将来的には800 ppm程度に達すると予測されています。しかし、将来の高CO₂環境下において、変動光に対する光合成制御機構がどのように働くのかについては、十分には明らかになっていませんでした。
そこで本研究では、光合成酵素Rubiscoを活性化するタンパク質「Rubisco activase」に着目しました。Rubisco activaseは、Rubiscoの活性状態を調節することで光合成を制御する重要因子として知られています。本研究では、Rubisco activaseを増やしたイネおよび減少させたイネを用い、変動光環境下における光合成応答や植物成長への影響を解析することで、将来の高CO₂環境下での作物生産性向上に向けた新たな光合成改良戦略の可能性を検証しました。
Rubisco activase量を変えても定常状態の光合成能力は大きく変わらない
研究グループはまず、Rubisco activase過剰発現体およびノックダウン体を作出し、光合成関連特性を解析しました。その結果、Rubisco activase量は大きく変化している一方で、Rubisco量や定常状態における光合成能力には大きな差が見られませんでした(図1A–D)。また、高CO₂条件では、定常状態の光合成はRuBP再生によって制限されることが示されました(図1C, D)。これらの結果は、高CO₂環境では単純なRubisco量だけではなく、光合成調節機構そのものが重要になることを示しています。
Rubisco activaseが光合成誘導を高速化
次に、低光から高光へ急激に光強度を変化させた際の光合成誘導を解析しました。その結果、Rubisco activase過剰発現体では、光強度増加後のRubisco活性化速度が大きく向上し、それに伴って光合成速度の立ち上がりも高速化されました(図2A, B, I, J)。一方、ノックダウン体では、光合成誘導が大きく遅延しました(図2A, B)。また、気孔コンダクタンスには大きな違いが見られなかったことから(図2C,D,G,H)、光合成誘導の高速化は主にRubisco活性制御によるものであることが示されました。
長時間の変動光条件でも高い光合成能力と水利用効率を維持
研究グループは、長時間の変動光条件下で光合成応答を解析しました。その結果、Rubisco activase過剰発現体では、高い光合成速度が長時間維持されました(図3)。一方、ノックダウン体では、変動光を繰り返すことで光合成能力が低下しました(図3)。
さらに、5時間積算光合成量および水利用効率(WUEi)を解析した結果、Rubisco activase過剰発現体では、変動光条件下で炭素固定量と水利用効率が向上していました(図4)。特に高CO₂条件では、その効果がより顕著でした(図4)。
Rubisco activaseが変動光によるPSI損傷を軽減
変動光環境では、光化学系I(PSI)が損傷を受けやすいことが知られています。本研究では、Rubisco activase過剰発現体では、変動光処理後もPSI活性が高く維持されていた一方で、ノックダウン体ではPSI損傷が顕著に増加しました(図5)。 さらに、変動光処理後のRubisco活性化状態、Rubisco最大カルボキシル化速度(Vcmax)、電子伝達速度(J)を解析した結果、Rubisco activase過剰発現体では高いRubisco活性状態および光合成能力が維持されていることが分かりました(図6)。これらの結果は、Rubisco activaseが効率的なCO₂固定を維持することで、過剰還元状態を抑え、PSI保護にも寄与していることを示しています。
将来の高CO₂環境でも効果を確認
現在、大気中CO₂濃度は増加を続けており、将来的には800 ppm程度に達すると予測されています。本研究では、この将来環境を模擬した高CO₂条件下でも解析を行いました。その結果、Rubisco activase過剰発現体では、高CO₂環境下でも光合成誘導が高速化され、長時間の変動光条件において高い光合成能力を維持しました(図2〜4)。 さらに、実際の水田環境で測定した自然光変動を再現した条件でも、高い光合成速度を維持し(図7)、積算炭素固定量および水利用効率が向上していました(図7D–G)。一方、ノックダウン体ではこれらの値が低下しました(図7)。これらの結果から、Rubisco activaseは将来の高CO₂環境下においても、自然環境に近い変動光条件で重要な役割を果たすことが明らかとなりました。
変動光環境下でイネの成長を促進
最後に、一定光条件および変動光条件下で40日間の栽培試験を行いました(図8A, B)。その結果、一定光条件では系統間で大きな差は見られなかった一方で、変動光条件ではRubisco activase過剰発現体で乾燥重量が有意に増加し、成長促進が確認されました(図8C, D)。逆に、ノックダウン体では成長が低下しました(図8D)。 これらの結果は、Rubisco activaseによる光合成誘導の高速化が、実際の変動光環境下における作物生産性向上につながることを示しています。
今後の展望
本研究は、Rubisco activaseが、将来の高CO₂環境下においても、変動光条件での光合成と成長を向上させる重要因子であることを示しました。
今後は、高温環境との複合ストレス条件や、より複雑な自然光変動条件における解析を進めることで、気候変動下でも高い生産性を維持できる作物開発への応用が期待されます。また、Rubisco activaseの改変は、水利用効率向上にも寄与することから、将来の気候変動環境に適応した高生産性作物の開発につながることが期待されます。
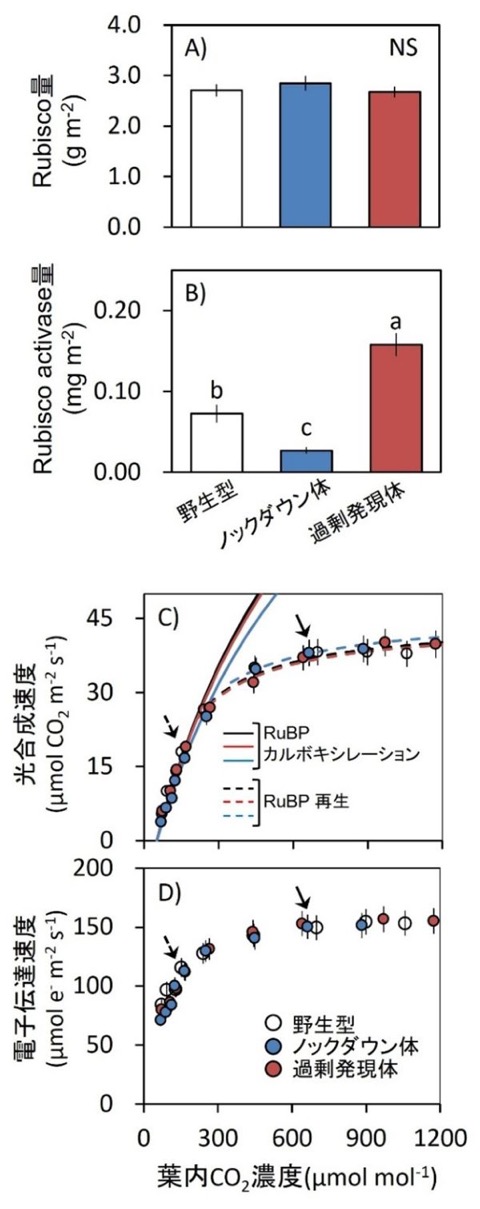 図1.光合成に関する生理特性
図1.光合成に関する生理特性
(A, B)高CO₂条件(800 µmol mol⁻¹)で栽培した野生型、Rubisco activase過剰発現体、およびRubisco activaseノックダウン体における、葉面積当たりのRubisco量(A)およびRubisco activase量(B)。
(C, D)25°C、1500 µmol photons m⁻² s⁻¹ 条件下における、光合成速度(A;C)および光化学系IIを介した電子伝達速度(ETR II;D)のCO₂応答曲線。C3光合成モデルを用いて、Rubisco律速条件(Ac:実線)とRuBP再生律速条件(Ar:点線)を推定した。矢印は大気CO₂濃度400または800 µmol mol⁻¹における値を示す。
白丸:野生型、赤丸:Rubisco activase過剰発現体、青丸:Rubisco activaseノックダウン体。データは平均値 ± SEM(野生型:n = 4、過剰発現体およびノックダウン体:n = 8)。異なるアルファベットは有意差あり、NSは有意差なしを示す(Tukey–Kramer HSD検定)。
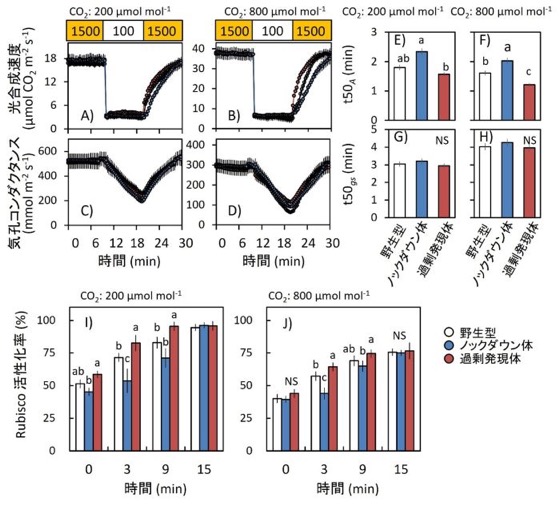 図2.短時間の変動光に対する光合成応答
図2.短時間の変動光に対する光合成応答
(A–D)野生型、Rubisco activase過剰発現体、およびRubisco activaseノックダウン体における、光強度増加後の光合成速度(A, B)および気孔コンダクタンス(C, D)の変化。測定はCO₂濃度200 µmol mol⁻¹(A, C)または800 µmol mol⁻¹(B, D)条件下で行った。植物は800 µmol mol⁻¹ CO₂条件で栽培した。葉はまず高光(1500 µmol photons m⁻² s⁻¹)下で30分間順化させて定常状態にした後、低光(100 µmol photons m⁻² s⁻¹)に12分間曝露し、その後再び高光(1500 µmol photons m⁻² s⁻¹)を照射した。ガス交換およびクロロフィル蛍光パラメータは30秒ごとに記録した。白丸:野生型、赤丸:Rubisco activase過剰発現体、青丸:Rubisco activaseノックダウン体。
(E–H)光強度増加後に最大値の50%へ到達するまでの時間(t50)を用いた光合成誘導速度解析。光合成速度t50A(E, F)および気孔コンダクタンスt50gs(G, H)について、CO₂濃度200 µmol mol⁻¹(E, G)または800 µmol mol⁻¹(F, H)条件下で解析した。
(I, J)光強度増加後のRubisco活性化状態の変化。CO₂濃度200 µmol mol⁻¹(I)または800 µmol mol⁻¹(J)条件下で解析した。Rubisco活性化状態は、高光(1500 µmol photons m⁻² s⁻¹)照射後0、3、9、15分で測定した。
白:野生型、赤:Rubisco activase過剰発現体、青:Rubisco activaseノックダウン体。データは平均値 ± SEM(野生型:n = 4、過剰発現体およびノックダウン体:n = 8)。異なるアルファベットは有意差あり、NSは有意差なしを示す(Tukey–Kramer HSD検定)。
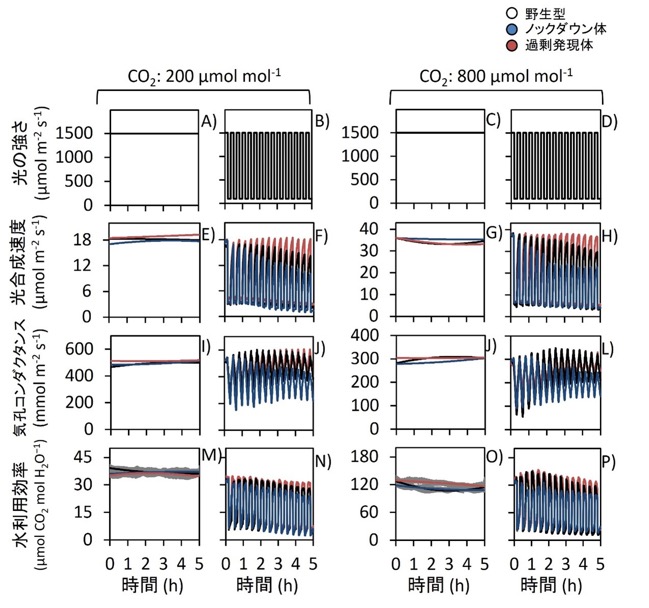 図3.長時間の変動光に対する光合成応答
図3.長時間の変動光に対する光合成応答
野生型、Rubisco activase過剰発現体、およびRubisco activaseノックダウン体における、長時間の一定光または変動光条件下での光合成応答。測定はCO₂濃度200 µmol mol⁻¹(A, B)または800 µmol mol⁻¹(C, D)条件下で行った。植物は800 µmol mol⁻¹ CO₂条件で栽培した。
光合成速度(E–H)、気孔コンダクタンス(I–L)、および光合成速度を気孔コンダクタンスで除して算出した水利用効率(WUEi;M–P)を、一定高光(1500 µmol photons m⁻² s⁻¹)または変動光条件[100 µmol photons m⁻² s⁻¹を10分間、1500 µmol photons m⁻² s⁻¹を10分間照射]下で5時間測定した。
葉はまず1500 µmol photons m⁻² s⁻¹条件下で30分以上順化し、定常状態の光合成に達した後、一定光または変動光条件下で長時間の光合成応答を解析した。
データは平均値 ± SEM(野生型:n = 4、過剰発現体およびノックダウン体:n = 8)。
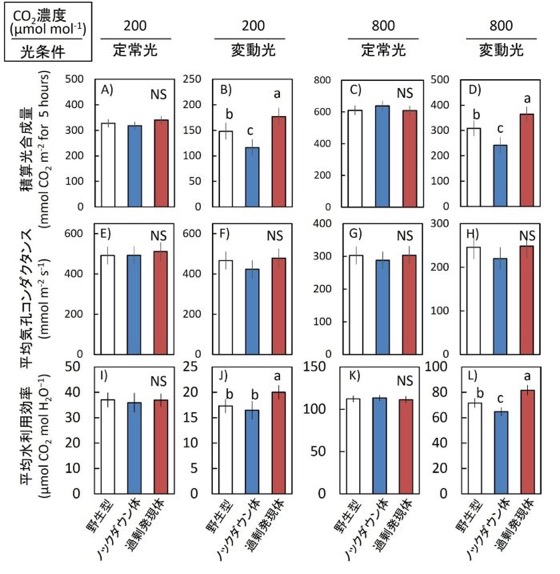 図4.5時間積算光合成量、平均気孔コンダクタンス、および平均水利用効率
図4.5時間積算光合成量、平均気孔コンダクタンス、および平均水利用効率
CO₂濃度200 µmol mol⁻¹または800 µmol mol⁻¹条件下における、一定高光または変動光条件での5時間積算光合成量、平均気孔コンダクタンス、および平均水利用効率[平均WUEi;光合成速度を気孔コンダクタンスで除して算出]を解析した。
光条件は図3と同一であり、解析には図3のデータを用いた。
データは平均値 ± SEM(野生型:n = 4、過剰発現体およびノックダウン体:n = 8)。異なるアルファベットは有意差あり、NSは有意差なしを示す(Tukey–Kramer HSD検定)。
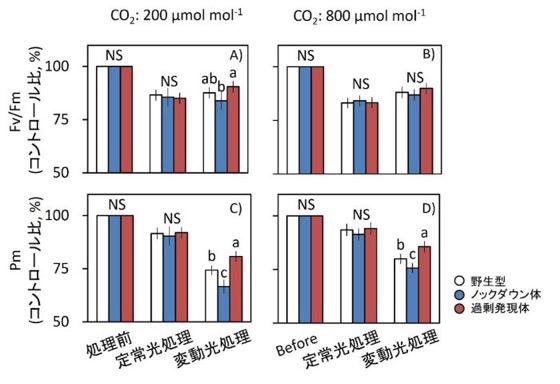 図5.長時間の一定光および変動光処理後の光阻害
図5.長時間の一定光および変動光処理後の光阻害
一定高光または変動光を5時間処理した後の、光化学系II(PSII)の最大量子収率(Fv/Fm)および光化学系I(PSI)の最大P700シグナル(Pm;P700の完全酸化レベル)を測定した。
測定は、植物を暗所で30分間保持した後に行った。光処理条件は図3と同一である。
値は、処理前の野生型を100%とした相対値(%, コントロール比)として示した。
データは平均値 ± SEM(野生型:n = 4、過剰発現体およびノックダウン体:n = 8)。異なるアルファベットは有意差あり、NSは有意差なしを示す(Tukey–Kramer HSD検定)。
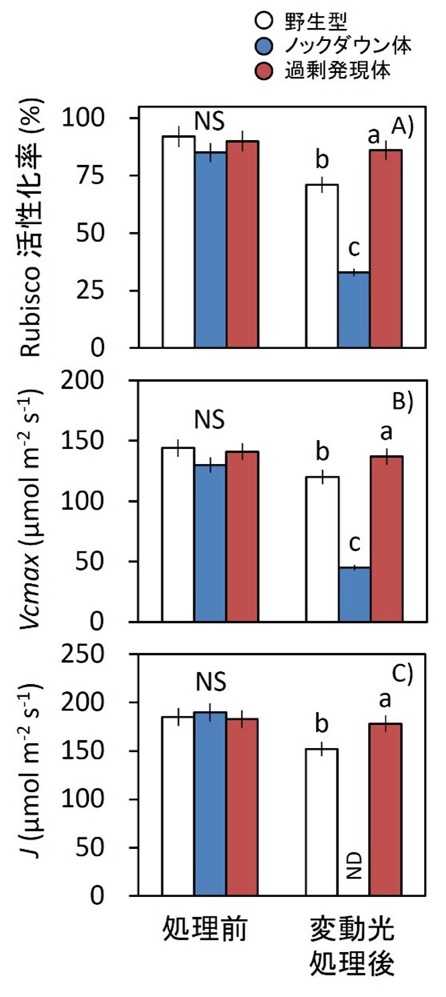 図6.長時間の変動光処理後の光合成パラメータ
図6.長時間の変動光処理後の光合成パラメータ
5時間の変動光処理前後における、Rubisco活性化率(A)、Rubisco最大カルボキシル化速度(Vcmax;B)、および電子伝達速度(J;C)を解析した。光処理条件は図3と同一である。C3光合成モデルを用いて、VcmaxとJを算出した。光合成速度のCO₂応答曲線の元データは補足図1に示した。
データは平均値 ± SEM(野生型:n = 4、過剰発現体およびノックダウン体:n = 8)。異なるアルファベットは有意差あり、NSは有意差なしを示す(Tukey–Kramer HSD検定)。NDは不検出を示す。
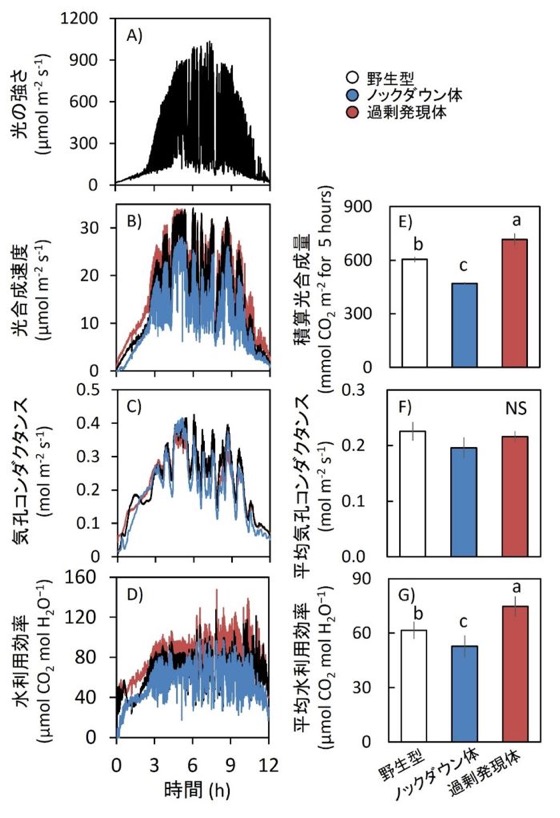 図7.自然光変動を模擬した条件下における光合成パラメータの経時変化
図7.自然光変動を模擬した条件下における光合成パラメータの経時変化
水田内のイネ群落で測定した自然光変動を、LI-6400XTシステムを用いて再現した(A)。光強度は実際に測定した自然光プロファイルに従って変動させ、ガス交換パラメータを10秒間隔で記録した。
CO₂濃度800 µmol mol⁻¹条件下で栽培した野生型、Rubisco activase過剰発現体、およびRubisco activaseノックダウン体について、光合成速度(B)、気孔コンダクタンス(C)、および光合成速度を気孔コンダクタンスで除して算出した水利用効率(WUEi;D)を測定した。
さらに、積算光合成量(E)、平均気孔コンダクタンス(F)、および平均水利用効率(WUEi;G)も算出した。
データは平均値 ± SEM(n = 4)。異なるアルファベットは系統間で有意差あり、NSは有意差なしを示す(Tukey–Kramer HSD検定)。NDは不検出を示す。
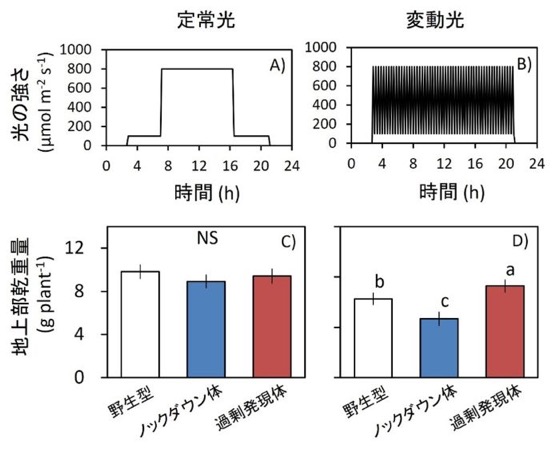 図8.一定光および変動光条件下における植物成長
図8.一定光および変動光条件下における植物成長
発芽後40日間、一定光条件(600 µmol photons m⁻² s⁻¹)で栽培した植物を用いた。
一定光処理における光条件を(A)に示した。植物は、朝と夕方に低光をそれぞれ4.5時間、日中に高光を9時間照射した。
変動光処理における光条件を(B)に示した。高光(10分間)と低光(10分間)を1日18時間交互に繰り返した。
一定光条件および変動光条件で40日間処理後の植物成長を、それぞれ(C)および(D)に示した。
データは平均値 ± SEM(野生型:n = 4、過剰発現体およびノックダウン体:n = 8)。異なるアルファベットは有意差あり、NSは有意差なしを示す(Tukey–Kramer HSD検定)。
発表者・研究者等情報
東京大学大学院農学生命科学研究科
稲垣 茉里香(研究当時:修士課程)
矢守 航 准教授
神戸大学
大学院農学研究科
深山 浩 教授
論文情報
雑誌名:Plant, Cell & Environment題名:Overexpression of Rubisco activase improves photosynthesis and plant growth in rice under fluctuating light under future high CO2 conditions
著者名:Wataru Yamori*, Marika Inagaki, Hiroshi Fukayama
DOI:https://doi.org/10.1111/pce.70617
URL:https://onlinelibrary.wiley.com/doi/10.1111/pce.70617
研究助成
本研究は、日本学術振興会 科学研究費助成事業 基盤研究(B)(課題番号:21H02171)、学術変革領域研究(A)「細胞質ゲノム制御」(課題番号:24H02277)、および科学技術振興機構(JST)ALCA-Next(課題番号:JPMJAN25D2)の支援を受けて実施されました。用語解説
(注1) Rubisco植物の光合成においてCO₂固定を担う中心酵素。地球上で最も多く存在するタンパク質。
(注2) Rubisco activase
Rubiscoを活性化するタンパク質。Rubiscoに結合した阻害物質をATP依存的に除去し、光合成を円滑に進める役割を持つ。
(注3) 変動光環境
自然界で見られるような、時間的に光強度が変化する環境。
(注4) 光合成誘導
暗い環境から明るい環境へ移行した際に、光合成速度が徐々に上昇する現象。
問い合わせ先
(研究内容については発表者にお問合せください)東京大学 大学院農学生命科学研究科 附属生態調和農学機構
准教授 矢守 航(やもり わたる)
TEL:070-6442-9511
E-mail:yamori@g.ecc.u-tokyo.ac.jp
東京大学大学院農学生命科学研究科・農学部
事務部 総務課総務チーム広報情報担当
TEL:03-5841-5484 FAX:03-5841-5028
E-mail:koho.a@gs.mail.u-tokyo.ac.jp