 理化学研究所 研究
理化学研究所 研究RNAを見分けてほどく、ヘリカーゼの分子機構
-タンパク質の柔軟な天然変性領域がRNA識別の鍵-
【産学連携対象 全学共通分野 Discovery Saga】
【Sagaキーワード】
環境変化/言語発達/原子核/磁気共鳴/閉じ込め/内部構造/スペクトル/磁場/芳香族/ポリペプチド/構造形成/アミド/らせん構造/蛍光スペクトル/物理化学/X線結晶構造解析/塩基配列/結晶構造解析/アミン/選択性/構造モデル/ドメイン構造/カリウム/電子顕微鏡/電磁波/構造予測/配列解析/神経発達/生体内/X線結晶構造/結晶構造/変異体/生合成/クライオ電子顕微鏡/アルギニン/転写制御因子/翻訳制御/アデノシン/がん遺伝子/悪性黒色腫/悪性脳腫瘍/浸潤/染色体/mRNA/ポリアミン/分子機構/翻訳調節/がん化/ATP/RNA/アミノ酸/がん細胞/グルタミン酸/コンフォメーション/トリプトファン/ヌクレオシド/ヘリックス/メラノーマ/遺伝子発現制御/核磁気共鳴/蛍光標識/高次構造/細胞骨格/細胞増殖/生体分子/創薬/相互作用解析/転写因子/転写制御/動的構造/発現制御/発現調節/分子認識/立体構造/ストレス
2025年8月28日
理化学研究所
バイオ産業情報化コンソーシアム
東京大学
概要
理化学研究所(理研)生命機能科学研究センター 生体分子動的構造研究チーム(研究当時)の嶋田 一夫 チームリーダー(研究当時、現生命医科学研究センター 生体分子動的構造研究チーム チームディレクター、バイオ産業情報化コンソーシアム(JBIC)特別顧問)、外山 侑樹 研究員(研究当時、現生命医科学研究センター 生体分子動的構造研究チーム 客員研究員、東京大学 大学院薬学系研究科 特任助教)、東京大学 大学院薬学系研究科 生命物理化学教室の竹内 恒 教授の共同研究チームは、メッセンジャーRNA(mRNA)[1]を認識し、その翻訳制御を担うRNAヘリカーゼ[2]の一種DEAD-Box RNA helicase 3 X-linked(DDX3X)[2]タンパク質が、特定のmRNAを選択的に見分ける分子機構の一端を明らかにしました。本研究成果は、DDX3Xが関わる疾患の発症機序の解明や、タンパク質とRNAの相互作用を標的とした新たな創薬技術の開発に貢献すると期待されます。
DDX3Xは、mRNAの高次構造[3]をほどくRNAヘリカーゼ活性を持ち、がんや神経発達症の発症などに関与していることが知られています。今回、共同研究チームは、溶液核磁気共鳴分光法(溶液NMR法)[4]を用いて、DDX3Xの天然変性領域(IDR)[5]が、mRNA中のグアニン4重鎖(GQ)構造[6]を選択的に認識し、ヘリカーゼ活性を担うコア領域がこのGQ構造をほどくことを明らかにしました。そして本研究は、DDX3Xがこれら二つの領域の協奏的な働きによってmRNAを選択的に認識し、その翻訳を促進する分子機構を提唱しました。
本研究は、科学雑誌『Nature Communications』オンライン版(8月28日付:日本時間8月28日)に掲載されました。
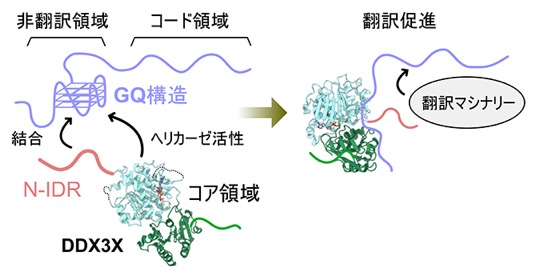
DDX3XがmRNAを選択的に認識し、翻訳制御を行う分子機構の模式図
背景
生物は遺伝子をコードするDNAからmRNAへの転写、mRNAからタンパク質の翻訳という過程を経ることで、さまざまな機能を持つタンパク質を合成し、生命活動を維持します。そして、これら一連の過程は、環境変化やストレスなど外的刺激に応じて、多様なメカニズムによって制御を受けています。mRNAからタンパク質の翻訳を制御するメカニズムの一つとして、mRNAの非翻訳領域[1]が複雑に折り畳まれた高次構造を形成し、タンパク質の翻訳速度や翻訳効率を抑制する機構が存在することが知られています。一方、細胞内には、mRNA非翻訳領域の高次構造をほどくことで翻訳を促進するタンパク質RNAヘリカーゼが存在します。このmRNAの非翻訳領域の高次構造形成とRNAヘリカーゼ活性のバランスにより、多くの重要な遺伝子の発現が精緻に制御を受けています。DDX3XはDEAD-Box RNAヘリカーゼの一種であり、細胞のがん化にも関与するRAC1遺伝子[7]やMITF遺伝子[8]など特定の遺伝子の翻訳促進をつかさどるタンパク質です。DDX3Xの活性が変調を受ける病因性変異は、髄芽腫(悪性脳腫瘍の一種)などのがんや、知的発達・言語発達の遅れなど神経発達症の要因となること、またDDX3Xを介したMITF遺伝子の翻訳調節はがん細胞の増殖、浸潤や転移といった病態の変化に関わることなどが知られています。このため、DDX3Xはがんや神経発達症を含む多様な疾病との関連が示唆される重要な創薬標的であると考えられています。従って、DDX3Xが、これら特定の遺伝子のmRNAをどのように認識し、その発現調節をつかさどるのかを分子レベルで明らかにすることは、DDX3Xの機能不全が関与する疾患に対して新規治療戦略を確立する上で重要です。
これまでにDDX3Xによって翻訳制御を受ける遺伝子群、およびそれらの遺伝子発現制御の生理的機能に着目した研究は盛んに行われてきたものの、DDX3Xが認識するmRNAには特定の配列モチーフ(共通して見られる特徴的な塩基配列)は見いだされておらず、その選択的な分子認識がいかにして達成されているのかは十分に明らかになっていませんでした。
研究手法と成果
今回、共同研究チームは、ヒト由来DDX3Xタンパク質を対象とし、まずはDDX3Xのどの領域が選択的なmRNA認識に関わっているかを調べました。DDX3XのRNAヘリカーゼ活性は、複数のαヘリックス[5]などで形成されたコア領域が担っています。一方、コア領域のN末端[9]とC末端[9]側には、特定の立体構造を形成しない天然変性領域(IDR)が存在します(図1A)。共同研究チームはこれらのN末端とC末端それぞれのIDRの機能に着目し、IDRを含まないDDX3X変異体タンパク質を作製してヘリカーゼ活性を測定しました。その結果、N末端側IDR(N-IDR)を持たないDDX3X変異体は正常型のDDX3Xと比較して活性が1/6倍程度にまで低下することが分かりました(図1B)。このことは、N-IDRがRNA基質の効率的な取り込みに重要な役割を担っていることを示します。次に、N-IDRがRNAに対する選択性を有しているのかを評価しました。DDX3Xが認識するRNAには特定の配列モチーフが見いだされていないことから、RNAの「配列」ではなく、「高次構造」を認識しているのではないかという仮説に基づき、N-IDRのみにしたDDX3Xタンパク質断片と、さまざまな高次構造を持つRNAとの相互作用を比較しました。N-IDRは構造的に柔軟な特性を有していることから、X線結晶構造解析[10]やクライオ電子顕微鏡構造解析[11]など、均一な構造を持つタンパク質に対する解析手法を適用するのが困難です。これに対し溶液NMR法は、柔軟なタンパク質分子であっても、NMRシグナル[4]の変化から部位特異的な相互作用の情報を得ることができます。そこで本研究では、溶液NMR法を活用することで、IDRによるRNAの認識機構を詳細に解析することとしました。
mRNAの非翻訳領域に含まれるとされる代表的な構造として、1本鎖RNA、2本鎖RNA[3]、ループ構造[3]を持つRNAがあります。まずこれらとの相互作用を解析したところ、N-IDRとの結合は観測されませんでした(図1C)。そこで解析対象を広げ、RNAの四つのグアニン同士が積み重なることで形成される特徴的な高次構造であるGQ構造を持つRNA(GQ RNA)との相互作用を解析したところ、N-IDRはGQ構造を持つRNAと強く相互作用することを見いだしました。GQ構造は、がん遺伝子の非翻訳領域などに含まれ、遺伝子の翻訳を抑制することが知られていることから、DDX3XのN-IDRはmRNAの非翻訳領域に形成されるGQ構造をマーカーとして選択的に認識することが示唆されました。
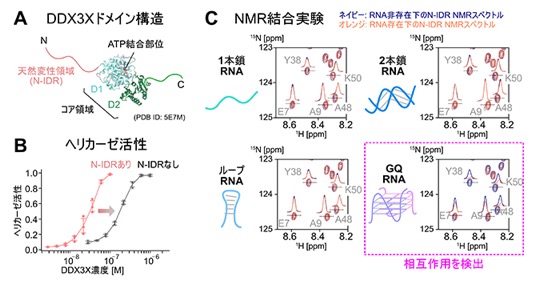
図1 DDX3Xタンパク質の構造とヘリカーゼ活性、およびN-IDRと各種RNAとの相互作用
A.DDX3Xドメイン構造。コア領域の立体構造はデータベース上で公開されているもの(PDB ID: 5E7M)を参照し、N、C末端の天然変性領域、コア領域を色分けして示した。
B.mRNAの高次構造モデル基質である2本鎖RNAに対するヘリカーゼ活性の測定結果。N-IDRを含む全長DDX3Xタンパク質を用いた結果を赤、N-IDRを持たないDDX3X変異体タンパク質の結果を黒でプロットした。
C.DDX3XのN-IDRの主鎖アミド基の15N-1H NMRシグナルを観測し、1本鎖RNA(左上)、2本鎖RNA(右上)、ループRNA(左下)、GQ RNA(右下)を添加した時のスペクトル変化を解析した。各スペクトルに添えられた記号は、観測したアミド基が属するアミノ酸残基(1文字記号で記す)がタンパク質中の何番目にあるかを示す(例:Y38→38番目のチロシン残基)。RNA添加前のスペクトルをネイビー、添加後のスペクトルをオレンジで重ね合わせて示した。1本鎖RNA、2本鎖RNA、ループRNAの三つはいずれもRNAの添加前後のスペクトルがほぼ重なっているのに対し、GQ RNA添加時には化学シフト値の変化やシグナル強度変化が顕著に観測されたことから、GQ RNAとより強く相互作用していることが分かった。
次に、GQ RNAとの相互作用によるN-IDRの部位特異的なNMRシグナルの変化を詳細に解析することで、N-IDRによるGQ認識機構を考察しました。解析の結果、N-IDRの配列の中には、正電荷を帯びたアミノ酸(アルギニン残基)が連なるクラスター領域が複数存在し、これらのクラスター領域がGQ RNAとの相互作用を担うこと、その認識には静電的な相互作用、疎水的な相互作用の両方が重要な役割を果たしていることを明らかにしました(図2A)。
GQ構造依存的なDDX3Xの認識が、実際の遺伝子のmRNA認識にも関連していることを示すため、DDX3Xによる翻訳制御を受けることが知られているRAC1遺伝子、ODC1遺伝子[12]、MITF遺伝子の非翻訳領域について、GQ構造予測プログラムを用いた配列解析(図2B)、およびNMRによるGQ構造形成の有無を評価したところ(図2C)、いずれの遺伝子にもGQ構造を形成する領域が存在することが分かりました。また、これらの遺伝子のmRNAからGQ構造形成領域を切り出したRNAフラグメントは、N-IDRと強い親和性で相互作用することも確認できました。これらの結果から、N-IDRはmRNAに含まれるGQ構造を介して特定遺伝子のmRNAを認識していることが支持されました(図2D)。
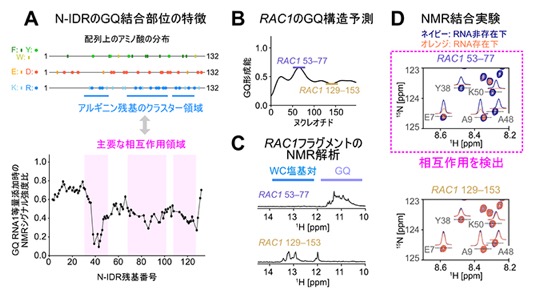
図2 N-IDRのGQ RNA認識部位とmRNA非翻訳領域のフラグメントRNAとの相互作用
A.(上)N末端の天然変性領域(N-IDR)を形成する132個のアミノ酸残基の配列上にある芳香族アミノ酸(Y:チロシン、Wトリプトファン)、負電荷を帯びたアミノ酸(E:グルタミン酸、D:アスパラギン酸)、および正電荷を帯びたアミノ酸(K:リジン、R:アルギニン)の分布を示した。(下)GQ RNAを1等量添加したときの、N-IDR各残基のシグナル強度比のプロット。顕著な強度減少を示した領域から、主要な相互作用領域を同定した。これらの相互作用領域は、配列上でアルギニン残基が集積したクラスター領域と一致することが分かった。
B.RAC1遺伝子のmRNA5'末端非翻訳領域の配列のGQ構造予測。構造予測はrG4 detectorソフトウェアによって行い、GQ構造を形成(53-77)、もしくは形成しない(129-153)領域のフラグメントを以後の解析に用いた。
C.RAC1の各フラグメントのイミノ領域(RNA塩基のNH部分)の1H NMRスペクトル。RAC1 53-77フラグメントではGQ構造に特徴的な化学シフトの領域にシグナルが観測され、GQ構造を形成していることが確認できた。なお「WC塩基対」は、相補塩基対で形成された典型的な核酸2本鎖構造(Watson-Crick型)を示す領域。
D.RAC1 53-77、もしくは129-153フラグメントを用いたN-IDRとのNMR相互作用解析の結果。RNA添加前のスペクトルをネイビー、添加後のスペクトルをオレンジで重ね合わせて示した。RAC1 53-77添加時に化学シフト値の変化やシグナル強度変化が顕著に観測されたことから、GQ構造を形成したフラグメントと相互作用していることが示された。
mRNAの非翻訳領域でGQ構造が形成されると、その翻訳が抑制されることが知られています。そこで共同研究チームは、DDX3Xはヘリカーゼ活性によってGQ構造をほどき、翻訳を促進する機能を持つのではないかと考えました。N-IDRとヘリカーゼ活性を担うコア領域の両方を持つ全長DDX3Xを用い、蛍光標識GQ RNAに対するヘリカーゼ活性を測定したところ、DDX3XはGQ構造をほどく活性を有していること、その活性効率はN-IDRに大きく依存していることが示されました(図3A)。以上の結果から、DDX3XはN-IDRによるGQ RNAの構造特異的な認識と、GQ RNAをほどくコア領域のヘリカーゼ活性が協奏的に働くことで、特定のmRNAを選択的に認識し、翻訳調節を効率的につかさどることを明らかにしました(図3B)。
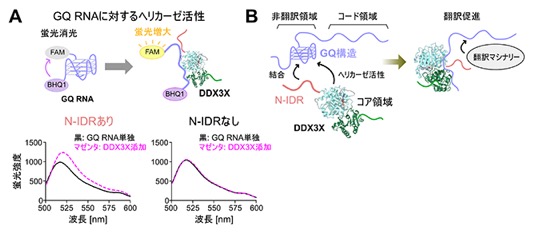
図3 DDX3XのGQ RNAに対するヘリカーゼ活性のアッセイと翻訳制御機構の模式図
A.(上)DDX3XのGQ RNAに対するヘリカーゼ活性の測定の模式図。二つの分子団の距離に応じて蛍光の強さが切り替わる仕組みを応用した。蛍光を発する蛍光団(FAM)と、その蛍光を打ち消す消光団(BHQ1)を付加したGQ RNAに対して、DDX3XがGQ構造をほどくと両者の距離が増大することで蛍光の増大が観測される。(下)N-IDRを含む全長DDX3Xタンパク質を用いた結果を左、N-IDRを持たないDDX3X変異体タンパク質の結果を右に示し、GQ RNA単独状態、DDX3X添加状態の蛍光スペクトルをそれぞれ黒、マゼンタで重ねて示した。変異体タンパク質は、ヘリカーゼ活性を担うコア領域を持っていても蛍光強度の変化が見られなかった。
B.本研究成果で得られたDDX3Xの分子機構の模式図。DDX3XのN-IDRがmRNAの非翻訳領域に形成されるGQ構造をマーカーとして選択的に認識し、ヘリカーゼ活性を担うコア領域がGQ構造をほどくことで翻訳を促進する。
今後の期待
本研究では、翻訳を促進する機能を持つRNAヘリカーゼDDX3XのN-IDRに着目し、さまざまな高次構造を有するRNAとの相互作用、およびそのRNA認識の選択性を、溶液NMR法を活用することで解析しました。本研究から明らかになったDDX3Xの分子機構は、DDX3Xが関与する遺伝子発現制御の分子レベルでの理解を深めるとともに、がんや神経発達症など、DDX3Xを標的とした創薬の基盤となる知見を供します。さらに、本研究で溶液NMR法を用いて明らかとなったIDRによるmRNA認識機構は、従来の「鍵と鍵穴」で例えられるような強固、かつ特異的な分子間相互作用ではなく、タンパク質分子の柔軟な性質を生かした弱く、かつ非特異的な分子間相互作用が集積することで機能する新たな認識様式であることが示されました。
IDRはヒトのプロテオーム(全タンパク質種)の大きな割合(おおよそ30%以上)を占め、機能的にも重要でありながら、特定の立体構造を示さないため創薬標的として極めて困難と考えられています。本研究の成果はIDRに対する新たな創薬戦略の指針を与えるものであり、特に、溶液NMR法から得られる分子・原子レベルでの知見は、既存の構造解析手法では解析が困難な、柔軟かつ動的なIDR-RNA相互作用に干渉する、新たな作用機序を持つ創薬技術の開発に貢献すると期待されます。
補足説明
1.メッセンジャーRNA(mRNA)、非翻訳領域DNAから転写され、細胞が合成するタンパク質のアミノ酸の並び方の情報を持つRNAをメッセンジャー(mRNA)と呼ぶ。mRNAは、タンパク質をコードする翻訳領域に加えて、5'、3'末端にはそれぞれタンパク質をコードしない非翻訳領域が存在する。一部の遺伝子産物のmRNAの5'末端の非翻訳領域は2本鎖やループ、GQ構造などさまざまな高次構造([3]参照)を形成することが知られており、これらの高次構造形成は翻訳マシナリーの結合や機能を阻害することで、翻訳を抑制的に制御する。RNAヘリカーゼ([2]参照)は、これらの高次構造をほどくことによって、特定の遺伝子の翻訳を促進する。
2.RNAヘリカーゼ、DEAD-Box RNA helicase 3 X-linked(DDX3X)
RNAヘリカーゼは、RNAのヘリックス(らせん)構造などの高次構造([3]参照)を解消する機能を持つ酵素。DEAD-Box RNAヘリカーゼは、活性部位にアスパラギン酸-グルタミン酸-アラニン-アスパラギン酸(D-E-A-D)から成る保存された配列モチーフを持つことを特徴とする、RNAの高次構造をATP依存的にほどくRNAヘリカーゼのファミリーであり、ヒトRNAのヘリカーゼにおいて最も大きなファミリーを構成する。DEAD-Box RNA helicase 3 X-linked(DDX3X)はX染色体上にその遺伝子が存在するDEAD-Box RNAヘリカーゼの一種である。
3.高次構造、2本鎖RNA、ループ構造
アデノシン(A)、シチジン(C)、グアノシン(G)、ウリジン(U)の4種類のリボヌクレオシドから構成されるRNAが、分子間もしくは分子内で相互作用することで形成される立体構造をRNAの高次構造と呼ぶ。RNAの2本鎖構造は、分子間でAとU、もしくはGとCとの間で相補的な塩基対を組むことで形成される高次構造である。DNAの二重らせん型構造と類似しているが、リボースのコンフォメーション(立体配座)が異なるA型のらせん構造を典型的に形成することを特徴とする。ループ構造はRNAに一般的に見られる高次構造の一つで、同じ分子内でRNA鎖が逆平行に向かい合い塩基対を組むことで形成される。
4.溶液核磁気共鳴分光法(溶液NMR法)、NMRシグナル
溶液中の化合物や生体分子などを対象に、強い磁場中に置くことで生じる原子核の共鳴現象を観測することで、その化学構造や運動性といった情報を原子レベルで解析できる分光法。磁場の中に置いた試料に電磁波を照射し、そこから放出される電磁波をNMRシグナルとして検出する。NMRはNuclear Magnetic Resonanceの略。
5.天然変性領域(IDR)、αヘリックス
タンパク質の立体構造は、アミノ酸がつながってできた「ひも」状の構造が、らせん状(αヘリックス)や平面状(βシート)などに折り畳まれて形成される。一方、固有の高次構造を形成しない領域は天然変性領域(IDR)と呼ばれ、ヒトのプロテオームの30%以上を占めるという試算もある。多くのタンパク質がIDRを有していると考えられているものの、IDRがタンパク質の生理的機能に果たす役割は明らかになっていないケースも多い。IDRはIntrinsically disordered regionの略。
6.グアニン4重鎖(GQ)構造
DNAやRNAにおいて、四つのグアニン塩基が同じ平面上で非ワトソン・クリック型塩基対であるフーグスティーン型塩基対を形成し、重なり合うことで形成される特徴的な高次構造。グアニン塩基の中央には、カリウムイオンなどの1価の陽イオンが配位することで安定化に寄与する。GQはG-quadruplexの略。
7.RAC1遺伝子
RAC1タンパク質をコードする遺伝子。RAC1は細胞増殖や細胞骨格の形成などの機能を担う、低分子量グアニンヌクレオチド結合タンパク質の一種。RAC1遺伝子の変異は、がん化を強く促進することでも知られている。RAC1はRAS-related C3 botulinus toxin substrate 1の略。
8.MITF遺伝子
小眼球症関連転写因子タンパク質をコードする遺伝子。MITFタンパク質はメラニン生合成の調節を担う転写制御因子である。MITFの発現量は皮膚がんの一種であるメラノーマ(悪性黒色腫)において、がんの転移能と相関していることが示されており、がんの薬剤耐性とも関連することが知られている。MITFはMicrophthalmia-associated transcription factorの略。
9.N末端、C末端
タンパク質はアミノ酸残基が直鎖状に連結したポリペプチドで構成されることから、その始まりと終わりがある。タンパク質の始まりとなる1番目の残基(またはその主鎖アミノ基)がN末端、終わりとなる最後の残基(またはその主鎖カルボキシル基)がC末端と呼ばれる。
10.X線結晶構造解析
物質の結晶をつくり、それにX線を照射して得られる回折データを解析することにより、物質の内部構造を調べる方法。
11.クライオ電子顕微鏡構造解析
タンパク質などの生体試料を観察するために開発された電子顕微鏡を用いた構造解析法。タンパク質などの試料を含んだ溶液を薄く展開し、液体エタン(-183~-160℃)中で急速凍結して試料をごく薄い氷の層に閉じ込めた上、液体窒素温度(-196℃)で電子顕微鏡により観察する。
12.ODC1遺伝子
オルニチンデカルボキシラーゼ1をコードする遺伝子。生体内におけるポリアミンの生合成に関与する酵素の一種。ODC1遺伝子をコードするmRNAは長く複雑な非翻訳領域を形成していることが知られている。また、神経芽腫などの一部のがんにおいてODC1の発現量が向上していることが報告されている。ODC1はornithine decarboxylase 1の略。
共同研究チーム
理化学研究所 生命機能科学研究センター生体分子動的構造研究チーム(研究当時)
チームリーダー(研究当時)嶋田 一夫(シマダ・イチオ)
(現 生命医科学研究センター 生体分子動的構造研究チーム チームディレクター、バイオ産業情報化コンソーシアム(JBIC)特別顧問、広島大学 副学長)
研究員(研究当時)外山 侑樹(トヤマ・ユウキ)
(現 生命医科学研究センター 生体分子動的構造研究チーム 客員研究員、東京大学 大学院薬学系研究科 特任助教)
東京大学 大学院薬学系研究科 生命物理化学教室
教授 竹内 恒(タケウチ・コウ)
研究支援
本研究は、理化学研究所運営費交付金(生命機能科学研究)で実施し、日本医療研究開発機構(AMED)「次世代治療・診断実現のための創薬基盤技術開発事業(RNA標的創薬技術開発)」による助成を受けて行われました。原論文情報
Yuki Toyama, Koh Takeuchi, Ichio Shimada, "Regulatory role of the N-terminal intrinsically disordered region of the DEAD-box RNA helicase DDX3X in selective RNA recognition",Nature Communications,10.1038/s41467-025-62806-7発表者
理化学研究所生命機能科学研究センター 生体分子動的構造研究チーム(研究当時)
チームリーダー(研究当時)嶋田 一夫(シマダ・イチオ)
(現生命医科学研究センター生体分子動的構造研究チーム チームディレクター、バイオ産業情報化コンソーシアム(JBIC)特別顧問)
研究員(研究当時)外山 侑樹(トヤマ・ユウキ)
(現 生命医科学研究センター 生体分子動的構造研究チーム 客員研究員、東京大学 大学院薬学系研究科 特任助教)
東京大学 大学院薬学系研究科 生命物理化学教室
教授 竹内 恒(タケウチ・コウ)
問い合わせ先
報道担当
理化学研究所 広報部 報道担当お問い合わせフォーム
バイオ産業情報化コンソーシアム 問い合わせ窓口
お問い合わせ | JBIC 一般社団法人バイオ産業情報化コンソーシアム
東京大学大学院薬学系研究科 庶務チーム
Tel: 03-5841-4702
Email: shomu@mol.f.u-tokyo.ac.jp